
载体构建的"利器"--Golden Gate

众所周知,基因功能研究,离不开分子生物学实验,而分子生物学实验又绕不开载体构建。依赖于限制性核酸内切酶的载体构建,其工作原理是分别对目的基因和载体DNA进行适当切割和修饰后,将二者连接在一起,再导入宿主细胞,实现目的基因在宿主细胞内的正确表达。目前常用的载体构建系统有:基于TypeII型限制性内切酶的载体构建系统、基于TypeIIs型限制性内切酶的Golden Gate载体构建系统。
与基于TypeII型限制性内切酶的载体构建系统不同,Golden Gate载体构建方法,可实现在单一管内同时进行酶切和连接反应,不需要进行凝胶纯化及分步的酶切、连接反应,大大缩短实验时间,并且重组克隆不会残留酶切位点,真正做到“无缝”拼接,重组效率高。今天小远就为大家重点介绍一下Golden Gate载体构建系统,希望大家看完本文后会对该系统有一个更全面的认识!
背景介绍
Golden Gate载体构建方法最早是由Engler等人在2008年提出的(Engler et al. 2008)。该方法是一种新型的酶切连接方法,与传统的酶切连接不同。传统的酶切连接方法采用标准的TypeII型限制性内切酶(例如EcoRI)切割DNA,这些限制酶通常识别4~8bp的回文序列,并在识别序列内部切割产生粘性末端或平末端。而Golden Gate是采用TypeIIs型限制酶(例如BsaI)切割DNA,这些酶是在识别序列以外剪切产生四个碱基的粘性末端(图1)。由于粘性末端不是识别序列的一部分,因此它们可以直接作为连接DNA的片段。在设计无误的情况下,识别位点不会出现在最终的质粒中,达到精确的无缝克隆,连接效率接近100%。此外,四个碱基的特异性末端能用来连接多个片段,从而在一个反应体系中实现多片段的克隆重组。
构建流程
(1)设计引物扩增目的片段
通过PCR向目的片段中引入IIs型限制酶(如BsaI、BsmBI等)的识别序列,识别序列加在引物的5'端,为了确保限制酶能稳定结合到DNA双链上并发挥切割作用,需在识别序列末端加上保护碱基。所加的保护碱基的数量和种类不是固定的,一般3bp的保护碱基就是足够的,但如果为了保险起见,也可以将保护碱基增加至6bp。另外,为了帮助大家更好的设计引物,我司官网专门开发了Golden Gate克隆引物设计网址(http://www.biorun.com/tools/)欢迎大家点击使用。
(2)酶切连接
利用TypeIIs限制性内切酶(BsaI)识别目的片段和载体上的酶切位点,并在酶切位点之外进行酶切。之后,在连接酶的作用下将目的片段和载体进行无缝连接,可实现单片段克隆重组(图1)或多片段重组(图2)。并且,在合适的实验条件和实验设计下,利用Golden Gate可以组装超过20个DNA片段。
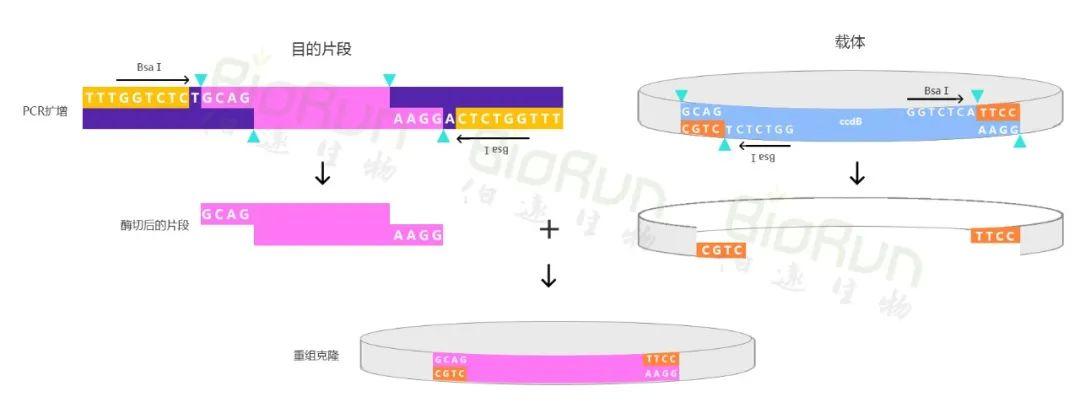

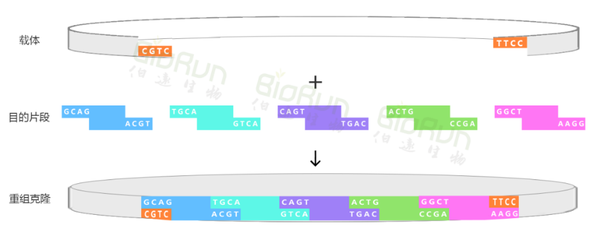

小远叨叨
就操作而言,Golden Gate克隆是最简单的方法之一,可在单一管内同时进行酶切和连接反应。并且,因为TypeIIs限制性内切酶剪切的特殊性,该方法的重组效率非常高。但是,在实际的操作中还是有一些注意事项需要提前了解,小远通过日常的实验经验以及阅读相关文献,总结了如下四点注意事项,一起来看看吧!
Golden Gate载体构建避坑指南
(1)DNA片段之间连接的粘性末端序列设计
4个碱基的粘性末端共有256种碱基排列方式,其中回文序列有16种,为了防止粘性末端自连而降低克隆效率,设计时应选用其他240种非回文序列(Engler et al., 2009)。
(2)选择合适的缓冲液
因为内切酶和连接酶同时存在于同一反应体系,所以缓冲液的选择尤为重要。在限制性核酸内切酶4种NEBuffer中,T4 DNA连接酶都具有有效活性(https://www.neb.com/)。因此能够推断,多数情况下反应体系可以考虑首选限制性内切酶的缓冲液。
(3)观察载体的酶切位点
构载体之前好好检查一下,质粒图谱上是否存在设计的酶切位点,存在几个。如果没有,要尽早更换酶切位点。为了避免不必要的消化,所使用的TypeIIs酶的酶切位点不能出现在需要连接的片段中。如果因为情况特殊出现的话,目的片段的剪切要换用其它的TypeIIs型限制酶,并且需要确保酶切后的粘性末端可以与载体进行组装,实验时需要对载体和目的片段分别进行酶切后再进行连接。
(4)控制反应体系中目的片段及载体浓度
在酶切含量一定的情况下,若目的片段及载体的浓度过高,可能会造成不完全酶切。
Golden Gate的应用
(1)合成生物学
随着合成生物学的发展,工程活性材料(ELMs)也出现了很多新的应用领域,利用合成生物学产生的材料具有突出特点,比如色泽、自我修复以及电导率等(Gilbert et al., 2019; Tang et al., 2021),此外,材料的生物可降解性,符合循环发展的理念。为了更好的推进合成生物学的发展,该领域的生物学家们利用Golden Gate克隆技术形成了模块化克隆策略。通过设计独一无二的限制酶切割位点帮助DNA片段按顺序连接,实现了多个DNA组分(启动子、基因、终止子等)在一个反应中准确连接。
例子1
2021年,Goosens等人在Komagataeibacteri 菌中构建了一个基于Golden Gate的标准化模块克隆系统Komagataeibacter Tool Kit(KTK)(图3),包括单基因系统和多基因系统。为了扩充该系统的范围,作者对不同的启动子进行表征,包括异源启动子和内源性启动子,筛选到J23104和PTac启动子是最强的异源启动子,PTtcA和PHxu是内源性启动子中最强的,同时作者还测试了不同的荧光报告蛋白在上述启动子下的表达强度(图4)。最后,作者利用KTK系统在Komagataeibacteri 中表达生产淀粉样纤维蛋白,作者将源自大肠杆菌淀粉样纤维蛋白生产系统导入Komagataeibacteri 菌内,并结合透射电子显微镜观察Komagataeibacteri 内生产淀粉样纤维蛋白的情况,因为在该菌中存在大量的纤维素,所以作者利用免疫标记来区分淀粉样纤维与纤维素,结果显示,在该菌中检测到有淀粉样纤维蛋白产生(图5),这也证实了该质粒在Komagataeibacteri 中的作用(Goosens et al., 2021)。
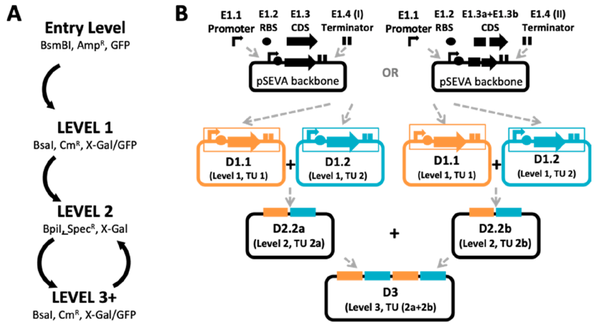

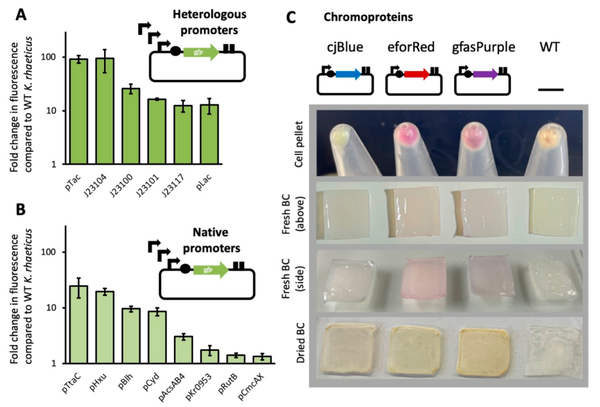

图4 用不同启动子驱动KTK系统(Goosens et al., 2021)。(A-B)通过流式细胞术测定的单个细胞的荧光,A图显示由6个不同的异源启动子分别驱动荧光蛋白表达的结果,B图显示由8个内源性启动子分别驱动荧光蛋白表达的结果。(C)不同的荧光报告蛋白(蓝色:cjBlue、红色:eforRed和紫色:gfasPurple)在J23104启动子下的表达强度。
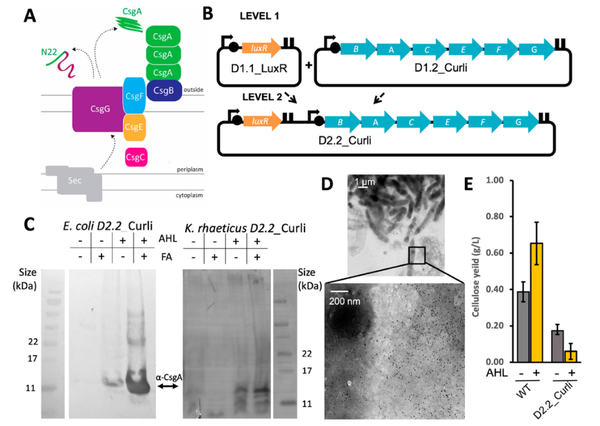

图5 在Komagataeibacteri中表达可诱导的Curli系统(Goosens et al., 2021)。(A)大肠杆菌Curli系统的示意图,其中淀粉样纤维蛋白由CsgA的聚合物组装而成,通过CsgB从核到细胞表面。这些蛋白质通过专门的VIII型分泌(T8S)系统(CsgE、CsgF、CsgG)被转移到外膜上,所有蛋白质都通过Sec系统被分泌到周质中。(B)载体示意图,D1.1_LuxR和D1.2_Curli被组装到一起,构建出了D2.2_Curli载体。(C)用D2.2_Curli质粒转化的大肠杆菌和Komagataeibacteri培养物中的蛋白质提取物的蛋白质印迹。(D)通过电子显微镜观察到的Komagataeibacteri内产生淀粉样纤维蛋白的情况。(E)野生型(WT)Komagataeibacteri和转化有D2.2_Curli的Komagataeibacteri的平均干表膜重量。
(2)基因组工程
在植物基因编辑中,CRISPR/Cas需要将筛选标记、Cas核酸酶编码基因和gRNA等多个表达元件一同转入植物,因此需要一个快速简便的克隆过程。基于Golden Gate克隆方法的模块化系统为多个DNA组分(启动子、基因、终止子等)的快速组装提供了解决方案。并且,这种载体构建的方法在CRISPR/Cas载体上组装单个/多个sgRNA中被运用到了极致。
例子1
2020年,Hahn等人在Golden Gate克隆方法模块化系统的基础上,开发了一个扩展的Golden Gate克隆工具包,其模块数量增至103个,适用于单子叶植物和双子叶植物基因编辑(Hahn et al., 2020)。扩展的Golden Gate克隆工具包中新增了编码FnCas12a、LbCas12a、Cms1核酸酶等元件、进行碱基替换(C-G到T-A和A-T到G-C)的碱基编辑器模块、具有PAM特异性的Cas9变异体(SaCas9、StCas9、ScCas9等)、Pol II和Pol III启动子以及gRNA骨架模块等。该工具包能够组装靶向单个以及多个靶标的CRISPR/Cas构建体,其gRNA在单个Pol III启动子下(图6)或使用多顺反子tRNA-sgRNA(图7)表达。tRNA-gRNA串联排列的多顺反子模块能够高效表达多个gRNA,并在植物中实现同时编辑多个基因组靶标位点,并且该过程无需PCR。作者通过在小麦原生质体中测试 tRNA-sgRNA CRISPR/Cas 载体,最终证实了该模块是有功能的(图8)。
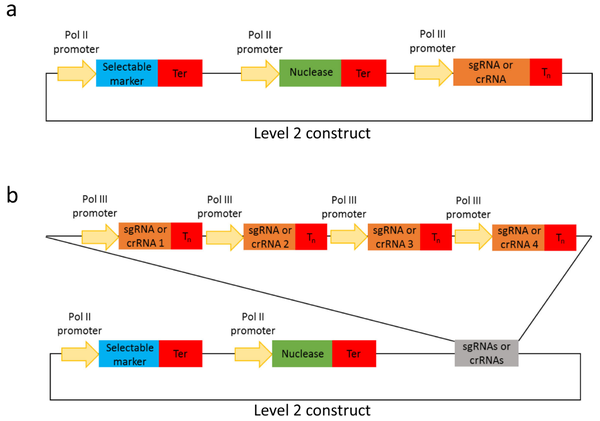

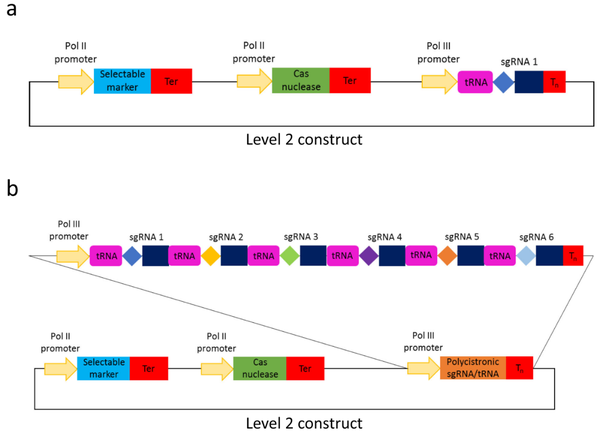



图8 在小麦原生质体中测试tRNA-sgRNA CRISPR/Cas载体(Hahn et al., 2020)。(a)三个小麦基因被编辑:基因1(同源染色体TraesCS1A02G338200,TraesCS1B02G350600和TraesCS1D02G340400),基因2(同源染色体TraesCS3A02G289300,TraesCS3B02G323900和TraesCS3D02G289100)和基因3(同源染色体TraesCS5A02G116500,TraesCS5B02G117800和TraesCS5D02G129600),红色箭头表示20bp sgRNA目标位点。(b)在小麦原生质体中共转携带三种SpCas9变异体(pFH23、pFH66 或 pFH67)之一的Leve 1载体和携带有6种sgRNA的多顺反子tRNA-sgRNA的Leve 1载体(pFH94)。通过 PCR 进行的基因分型结果显示了CRISPR/Cas诱导的片段缺失的情况。
例子2
2020年,张扬等人基于Golden Gate载体构建方法,建立了一种简单快速高效且无PCR扩增的双靶点CRISPR/Cas9载体构建方法(张扬,2020)。与传统的构建双靶点CRISPR/Cas9载体的方法不同,在该方法中,只需将设计好的2个sgRNA和1个外源片段用T4 DNA连接酶构建到CRISPR/Cas9载体上即可(图9),这大大降低了构建双靶点CRISPR/Cas9载体时操作的复杂性和实验时长。另外,由于该方法构建的CRISPR/Cas9载体不需要经过PCR反应过程,因此在很大程度上降低了外源片段的突变概率。作者使用该方法对10个已报道的水稻基因构建了20个双靶点CRISPR/Cas9载体,最终获得的阳性率为100%,测序结果显示无突变。
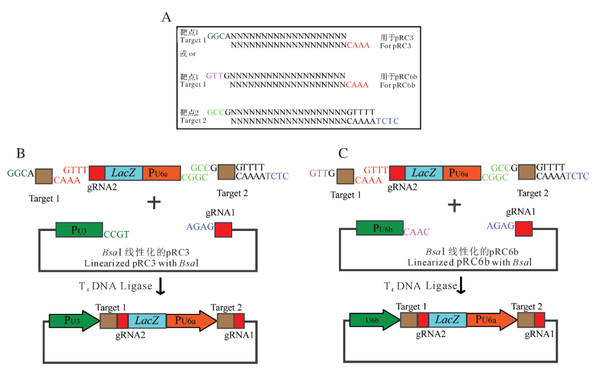

图9 双靶点CRISPR/Cas9载体构建流程图(张扬,2020)。(A)靶点设计示意图。如果靶点1的第1个碱基是A,则在正向靶点引物的5′端加GGC,如果靶点1的第1个碱基是G,则在正向靶点引物的5′端加GTT,反向靶点引物的5′端都是加AAAC;第2个靶点正向靶点引物的5′端加GCC,3′端加GTTTT,反向靶点引物的5′端加CTCTAAAAC。(B-C)载体构建流程图。2对设计好的靶点引物分别变性退火后,与gRNA2-LacZ-PU6a一起连入BsaⅠ线性化的pRC3或pRC6b,即可转化大肠杆菌感受态细胞。
小远叨叨
文章至此就告一段落了,本文主要为大家讲解了Golden Gate的背景优势、构建流程以及应用,相信大家通过阅读本文会对Golden Gate克隆技术有一个新的认识,今后进行无缝克隆的时候可以尝试用一下Golden Gate,真的会惊艳到你!另外,为了帮助大家更好的理解Golden Gate克隆技术的原理,小远专门翻译了一条视频帮助大家理解,大家一定要点击文末彩蛋观看视频哦~
References:
Engler C, Kandzia R, Marillonnet S. A one pot, one step, precision cloning method with high throughput capability. PLoS One. 2008, 3(11): e3647.
Engler C, Gruetzner R, Kandzia R, et al. Golden gate shuffling: a one-pot DNA shuffling method based on type IIS restriction enzymes. PLoS One, 2009, 4(5): e5553.
Gilbert C, Ellis T. Biological Engineered Living Materials: Growing Functional Materials with Genetically Programmable Properties. ACS Synth Biol. 2019, 8(1): 1-15.
Goosens VJ, Walker KT, Aragon SM, et al. Komagataeibacter Tool Kit (KTK): A Modular Cloning System for Multigene Constructs and Programmed Protein Secretion from Cellulose Producing Bacteria. ACS Synth Biol. 2021, 10(12): 3422-3434.
Hahn F, Korolev A, Sanjurjo Loures L, et al. A modular cloning toolkit for genome editing in plants. BMC Plant Biol. 2020, 20(1):179.
Tang T, An B, Huang Y, et al. Materials Design by Synthetic Biology. Nat.Rev. Mater. 2021, 6: 332.
张扬, 黄维峰, 周菲, 等. 1种简单快速高效的双靶点CRISPR/Cas9载体构建方法.华中农业大学学报. 2020, 39(03): 9-18.
NO.1好文推荐Historical articles
NO.2技术服务Technical service
载体构建
多重基因编辑全套服务
高通量基因编辑检测
突变体库靶标设计及完整解决方案